Классификация видов мышечной ткани. Поперечнополосатая мышечная ткань
Мышечные ткани – это специализированные ткани, основной функцией которых является сокращение. Благодаря им обеспечиваются все двигательные процессы в организме (гемоциркуляция в сосудах, ритмическая деятельность миокарда, перистальтика пищеварительного тракта и другие, а также перемещение организма в пространстве). Сокращение структурных элементов мышечных тканей осуществляется с помощью специальных органелл – миофибрилл – и является результатом взаимодействия молекул сократительных белков.
Существуют две классификации мышечных тканей – морфофункциональная и генетическая. Согласно первой классификации мышечные ткани делят на две группы: 1) гладкая (неисчерченная) мышечная ткань, которая характеризуется тем, что содержит миофибриллы, не имеющие поперечной исчерченности; 2) поперечнополосатая (исчерченная) мышечная ткань, миофибриллы которой образуют поперечную исчерченность. В свою очередь, она подразделяется на скелетную и сердечную . Согласно генетической классификации (по происхождению), мышечные ткани делят на 5 типов: 1) мезенхимные (развиваются из мезенхимы, находятся во внутренних органах и сосудах); 2) эпидермальные (развиваются из кожной эктодермы, включают немышечные сокращающиеся клетки – миоэпителиальные клетки потовых, молочных, слюнных и слезных желез); 3) нейральные (развиваются из нервной трубки, к ним принадлежат гладкие миоциты мышц радужной оболочки глаза); 4) соматические (развиваются из миотомов мезодермы и образуют скелетную мышечную ткань); 5) целомические (развиваются из висцерального листка спланхнотома и образуют сердечную мышечную ткань). Первые три типа относятся к гладким мышечным тканям, остальные – к поперечнополосатым. К общим структурным признакам, характерным для мышечных тканей, следует отнести наличие: 1)специальных органелл – миофибрилл, благодаря взаимодействию их сократительных белков, осуществляется сокращение; 2)развитого трофического аппарата, обеспечивающего выполнение сократительной функции – митохондрий, гладкой эндоплазматической сети, включений гликогена и миоглобина; 3)развитого опорного аппарата в виде двуслойной оболочки с окружающей ее сетью волокон соединительной ткани.
Гладкая мышечная ткань
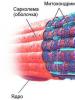
Гладкая мышечная ткань мезенхимного происхождения располагается в стенке внутренних органов и сосудов. Структурной единицей ее является гладкий миоцит . Это клетка веретеновидной, иногда отростчатой формы (матка, эндокард, аорта), длиной 20-500 мкм, с центрально расположенным ядром (рис. 7-1). Цитолемма гладкого миоцита образует многочисленные впячивания – кавеолы (мелкие пузырьки). Снаружи цитолемму покрывает тонкая базальная мембрана. В базальной мембране каждого миоцита есть отверстия, где клетки контактируют друг с другом при помощи нексусов, осуществляющих метаболические связи.
Органеллы общего значения – комплекс Гольджи, митохондрии, свободные рибосомы, саркоплазматическая сеть – локализуются в основном около полюсов ядра. Наиболее развитыми и многочисленными из них являются митохондрии . Саркоплазматическая сеть участвует в синтезе гликозаминогликанов и белковых молекул, из которых осуществляется сборка компонентов базальной мембраны, волокон, аморфного вещества, окружающих клетки. Синтетическая способность дефинитивных миоцитов снижается. Длинные узкие трубочки гладкой саркоплазматической сети, примыкают к кавеолам и вместе с ними служат для депонирования ионов кальция.
Специальные органеллы видны в виде нитей, ориентированных преимущественно вдоль длинной оси клетки и не имеющих поперечной исчерченности. В цитоплазме миоцитов стабильно выявляются только тонкие нити – миофиламенты, состоящие из белка актина. Они прикрепляются на внутренней стороне цитолеммы, образуя плотные тельца, состоящие из белка актинина. При изменении мембранного потенциала клетки ионы кальция, поступающие из депо, активируют сборку миозиновых (более толстых) нитей и их взаимодействие с актиновыми. По мере образования актин-миозиновых мостиков происходит смещение актиновых миофиламентов навстречу друг другу, тяга передается на цитолемму, и клетка укорачивается. При уменьшении содержания кальция миозин теряет сродство к актину. В результате начинается расслабление миоцита и разборка миозиновых нитей. Сокращение медленное, тоническое.
Рис. 7-1. Гладко-мышечная клет-ка.
1. Митохондрии.
2. Базальная мембрана.
3. Плотные тельца.
4. Зона щелевидных контактов.
5. Актиновые миофиламенты.
6. Ядро.
7. Кавеолы.
(По Lentz T. L. 1971).
Иннервация гладкой мышечной ткани осуществляется вегетативной нервной системой – симпатическими и парасимпатическими нервными волокнами, терминали которых формируют варикозные расширения на гладкомышечных клетках. Гладкие миоциты функционируют не изолированно, а клеточными комплексами. Клетки контактируют друг с другом при помощи нексусов. Последние способствуют проведению возбуждения от клетки к клетке, охватывая сразу группу миоцитов. В составе комплексов есть также миоциты-пейсмекеры, которые сами генерируют потенциал действия и передают его соседним клеткам.
Вокруг каждого гладкого миоцита из ретикулярных, эластических и коллагеновых волокон образуется сетка – эндомизий . Группы из 10-12 клеток объединяются в мышечные пласты, окруженные соединительной тканью с кровеносными сосудами и нервами, называемой перимизием . В органах пучки мышечных клеток формируют слои мышечной ткани. Совокупность пучков образует мышцу, которая окружена более толстой прослойкой соединительной ткани – эпимизием . При повышенной функциональной нагрузке гладкие миоциты гипертрофируются, как, например, в матке во время беременности, проявляя высокую способность к физиологической регенерации. При репаративной регенерации восстановление возможно за счет деления малодифференцированных миоцитов, которые находятся в составе мышечных комплексов, а также из адвентициальных клеток и миофибробластов.
По морфологическим признакам выделяют три группы мышц:
1) поперечно-полосатые мышцы (скелетные мышцы);
2) гладкие мышцы;
3) сердечную мышцу (или миокард).
Функции поперечно-полосатых мышц:
1) двигательная (динамическая и статическая);
2) обеспечения дыхания;
3) мимическая;
4) рецепторная;
5) депонирующая;
6) терморегуляторная.
Функции гладких мышц:
1) поддержание давления в полых органах;
2) регуляция давления в кровеносных сосудах;
3) опорожнение полых органов и продвижение их содержимого.
Функция сердечной мышцы – насосная, обеспечение движения крови по сосудам.
1) возбудимость (ниже, чем в нервном волокне, что объясняется низкой величиной мембранного потенциала);
2) низкая проводимость, порядка 10–13 м/с;
3) рефрактерность (занимает по времени больший отрезок, чем у нервного волокна);
4) лабильность;
5) сократимость (способность укорачиваться или развивать напряжение).
Различают два вида сокращения:
а) изотоническое сокращение (изменяется длина, тонус не меняется);
б) изометрическое сокращение (изменяется тонус без изменения длины волокна). Различают одиночные и титанические сокращения. Одиночные сокращения возникают при действии одиночного раздражения, а титанические возникают в ответ на серию нервных импульсов;
6) эластичность (способность развивать напряжение при растягивании).
Физиологические особенности гладких мышц.
Гладкие мышцы имеют те же физиологические свойства, что и скелетные мышцы, но имеют и свои особенности:
1) нестабильный мембранный потенциал, который поддерживает мышцы в состоянии постоянного частичного сокращения – тонуса;
2) самопроизвольную автоматическую активность;
3) сокращение в ответ на растяжение;
4) пластичность (уменьшение растяжения при увеличении растяжения);
5) высокую чувствительность к химическим веществам.
Физиологической особенностью сердечной мышцы является ее автоматизм . Возбуждение возникает периодически под влиянием процессов, протекающих в самой мышце. Способностью к автоматизму обладают определенные атипические мышечные участки миокарда, бедные миофибриллами и богатые саркоплазмой.
2. Механизмы мышечного сокращения
Электрохимический этап мышечного сокращения.
1. Генерация потенциала действия. Передача возбуждения на мышечное волокно происходит с помощью ацетилхолина. Взаимодействие ацетилхолина (АХ) с холинорецепторами приводит к их активации и появлению потенциала действия, что является первым этапом мышечного сокращения.
2. Распространение потенциала действия. Потенциал действия распространяется внутрь мышечного волокна по поперечной системе трубочек, которая является связывающим звеном между поверхностной мембраной и сократительным аппаратом мышечного волокна.
3. Электрическая стимуляция места контакта приводит к активации фермента и образованию инозилтрифосфата, который активирует кальциевые каналы мембран, что приводит к выходу ионов Ca и повышению их внутриклеточной концентрации.
Хемомеханический этап мышечного сокращения.
Теория хемомеханического этапа мышечного сокращения была разработана О. Хаксли в 1954 г. и дополнена в 1963 г. М. Девисом. Основные положения этой теории:
1) ионы Ca запускают механизм мышечного сокращения;
2) за счет ионов Ca происходит скольжение тонких актиновых нитей по отношению к миозиновым.
В покое, когда ионов Ca мало, скольжения не происходит, потому что этому препятствуют молекулы тропонина и отрицательно заряды АТФ, АТФ-азы и АДФ. Повышенная концентрация ионов Ca происходит за счет поступления его из межфибриллярного пространства. При этом происходит ряд реакций с участием ионов Ca:
1) Ca2+ реагирует с трипонином;
2) Ca2+ активирует АТФ-азу;
3) Ca2+ снимает заряды с АДФ, АТФ, АТФ-азы.
Взаимодействие ионов Ca с тропонином приводит к изменению расположения последнего на актиновой нити, открываются активные центры тонкой протофибриллы. За счет них формируются поперечные мостики между актином и миозином, которые перемещают актиновую нить в промежутки между миозиновой нитью. При перемещении актиновой нити относительно миозиновой происходит сокращение мышечной ткани.
Итак, главную роль в механизме мышечного сокращения играют белок тропонин, который закрывает активные центры тонкой протофибриллы и ионы Ca.
Физиология скелетных и гладких мышц
Лекция 5
У позвоночных и человека три вида мышц : поперечнополосатые мышцы скелета, поперечнополосатая мышца сердца – миокард и гладкие мышцы, образуюцие стенки полых внутренних органов и сосудов.
Анатомической и функциональной единицей скелетных мышц является нейромоторная единица - двигательный нейрон и иннервируемая им группа мышечных волокон. Импульсы, посылаемые мотонейроном, приводят в действие все образующие ее мышечные волокна.
Скелетные мышцы состоят из большого количества мышечных волокон. Волокно поперечнополосатой мышцы имеет вытянутую форму, диаметр его от 10 до 100 мкм, длина волокна от нескольких сантиметров до 10-12 см. Мышечная клетка окружена тонкой мембраной – сарколеммой , содержит саркоплазму (протоплазму) и многочисленные ядра . Сократительной частью мышечного волокна являются длинные мышечные нити – миофибриллы , состоящие в основном из актина, проходящие внутри волокна от одного конца до другого, имеющие поперечную исчерченность. Миозин в гладких мышечных клетках находится в дисперсном состоянии, но содержит много белка, играющего важную роль в поддержании длительного тонического сокращения.
В период относительного покоя скелетные мышцы полностью не расслабляются и сохраняют умеренную степень напряжения, т.е. мышечный тонус .
Основные функции мышечной ткани:
1)двигательная – обеспечение движения
2)статическая – обеспечение фиксации, в том числе и в определенной позе
3)рецепторная – в мышцах имеются рецепторы, позволяющие воспринимать собственные движения
4)депонирующая – в мышцах запасаются вода и некоторые питательные вещества.
Физиологические свойства скелетных мышц:
Возбудимость . Ниже, чем возбудимость нервной ткани. Возбуждение распространяется вдоль мышечного волокна.
Проводимость . Меньше проводимости нервной ткани.
Рефрактерный период мышечной ткани более продолжителен, чем нервной ткани.
Лабильность мышечной ткани значительно ниже, чем нервной.
Сократимость – способность мышечного волокна изменять свою длину и степень напряжения в ответ на раздражение пороговой силы.
При изотоническом сокращении изменяется длина мышечного волокна без изменения тонуса. При изометрическом сокращении возрастает напряжение мышечного волокна без изменения его длины.
В зависимости от условий стимуляции и функционального состояния мышцы может возникнуть одиночное, слитное (тетаническое) сокращение или контрактура мышцы.
Одиночное мышечное сокращение. При раздражении мышцы одиночным импульсом тока возникает одиночное мышечное сокращение.
Амплитуда одиночного сокращения мышцы зависит от количества сократившихся в этот момент миофибрилл. Возбудимость отдельных групп волокон различна, поэтому пороговая сила тока вызывает сокращение лишь наиболее возбудимых мышечных волокон. Амплитуда такого сокращения минимальна. При увеличении силы раздражающего тока в процесс возбуждения вовлекаются и менее возбудимые группы мышечных волокон; амплитуда сокращений суммируется и растет до тех пор, пока в мышце не останется волокон, не охваченных процессом возбуждения. В этом случае регистрируется максимальная амплитуда сокращения, которая не увеличивается, несмотря на дальнейшее нарастание силы раздражающего тока.
Тетаническое сокращение . В естественных условиях к мышечным волокнам поступают не одиночные, а ряд нервных импульсов, на которые мышца отвечает длительным, тетаническим сокращением, или тетанусом . К тетаническому сокращению способны только скелетные мышцы. Гладкие мышцы и поперечнополосатая мышца сердца не способны к тетаническому сокращению из-за продолжительного рефрактерного периода.
Тетанус возникает вследствие суммации одиночных мышечных сокращений. Чтобы возник тетанус, необходимо действие повторных раздражений (или нервных импульсов) на мышцу еще до того, как закончится ее одиночное сокращение.
Если раздражающие импульсы сближены и каждый из них приходится на тот момент, когда мышца только начала расслабляться, но не успела еще полностью расслабиться, то возникает зубчатый тип сокращения (зубчатый тетанус ).
Если раздражающие импульсы сближены настолько, что каждый последующий приходится на время, когда мышца еще не успела перейти к расслаблению от предыдущего раздражения, то есть происходит на высоте ее сокращения, то возникает длительное непрерывное сокращение, получившее название гладкого тетануса .
Гладкий тетанус – нормальное рабочее состояние скелетных мышц обусловливается поступлением из ЦНС нервных импульсов с частотой 40-50 в 1с.
Зубчатый тетанус возникает при частоте нервных импульсов до 30 в 1с. Если мышца получает 10-20 нервных импульсов в 1с, то она находится в состоянии мышечного тонуса , т.е. умеренной степени напряжения.
Утомление мышц . При длительном ритмическом раздражении в мышце развивается утомление. Признаками его являются снижение амплитуды сокращений, увеличение их латентных периодов, удлинение фазы расслабления и, наконец, отсутствие сокращений при продолжающемся раздражении.
Еще одна разновидность длительного сокращения мышц - контрактура . Она продолжается и при снятии раздражителя. Контрактура мышцы наступает при нарушении обмена веществ или изменении свойств сократительных белков мышечной ткани. Причинами контрактуры могут быть отравление некоторыми ядами и лекарственными средствами, нарушение обмена веществ, повышение температуры тела и другие факторы, приводящие к необратимым изменениям белков мышечной ткани.
Мышечные ткани - это ткани, отличающиеся по структуре и происхождению, но имеют общую способность к сокращению. Состоят из миоцитов - клеток, которые могут воспринимать нервные импульсы и отвечать на них сокращением.
Свойства и виды мышечной ткани
Морфологические признаки:
- Вытянутая форма миоцитов;
- продольно размещены миофибриллы и миофиламенты;
- митохондрии находятся вблизи сократительных элементов;
- присутствуют полисахариды, липиды и миоглобин.
Свойства мышечной ткани:
- Сократимость;
- возбудимость;
- проводимость;
- растяжимость;
- эластичность.
Выделяют следующие виды мышечной ткани в зависимости от морфофункциональных особенностей:
- Поперечнополосатая: скелетная, сердечная.
- Гладкая.
Гистогенетическая классификация делит мышечные ткани на пять видов в зависимости от эмбрионального источника:
- Мезенхимные - десмальный зачаток;
- эпидермальные - кожная эктодерма;
- нейральные - нервная пластинка;
- целомические - спланхнотомы;
- соматические - миотом.
Из 1-3 видов развиваются гладкомышечные ткани, 4, 5 дают поперечнополосатые мышцы.
Строение и функции гладкой мышечной ткани
Cостоит из отдельных мелких веретеновидных клеток. Эти клетки имеют одно ядро и тонкие миофибриллы, которые тянутся от одного конца клетки к другому. Гладкие мышечные клетки объединяются в пучки, состоящие из 10-12 клеток. Это объединение возникает благодаря особенностям иннервации гладкой мускулатуры и облегчает прохождение нервного импульса на всю группу гладких мышечных клеток. Сокращается гладкая мышечная ткань ритмично, медленно и на протяжении длительного времени, способна при этом развивать большую силу без значительных затрат энергии и без утомления.
У низших многоклеточных животных из гладкой мышечной ткани состоят все мышцы, тогда как у позвоночных животных она входит в состав внутренних органов (кроме сердца).
Сокращения этих мышц не зависят от воли человека, т. е. происходят непроизвольно.
Функции гладкой мышечной ткани:
- Поддерживание стабильного давления в полых органах;
- регуляция уровня кровяного давления;
- перистальтика пищеварительного тракта, перемещения по нему содержимого;
- опорожнение мочевого пузыря.
Строение и функции скелетной мышечной ткани

Cостоит из длинных и толстых волокон длиной 10-12 см. Скелетная мускулатура характеризуется произвольным сокращением (в ответ на импульсы, идущие из коры головного мозга). Скорость ее сокращения в 10-25 раз выше, чем в гладкой мышечной ткани.
Мышечное волокно поперечнополосатой ткани покрыто оболочкой - сарколеммой. Под оболочкой находится цитоплазма с большим количеством ядер, расположенных по периферии цитоплазмы, и сократительными нитями - миофибриллами. Состоит миофибрилла из последовательно чередующихся темных и светлых участков (дисков), обладающих разным коэффициентом преломления света. С помощью электронного микроскопа установлено, что миофибрилла состоит из протофибрилл. Тонкие протофибриллы построены из белка - актина, аболее толстые - из миозина.
При сокращении волокон происходит возбуждение сократимых белков, тонкие протофибриллы скользят по толстым. Актин реагирует с миозином, и возникает единая актомиозиновая система.
Функции скелетной мышечной ткани:
- Динамическая - перемещение в пространстве;
- статическая - поддержание определенной позиции частей тела;
- рецепторная - проприорецепторы, воспринимающие раздражение;
- депонирующая - жидкость, минералы, кислород, питательные вещества;
- терморегуляция - расслабление мышц при повышении температуры для расширения сосудов;
- мимика - для передачи эмоций.
Строение и функции сердечной мышечной ткани
 Сердечная мышечная ткань
Сердечная мышечная ткань
Миокард построен из сердечной мышечной и соединительной ткани, с сосудами и нервами. Мышечная ткань относится к поперечнополосатой мускулатуре, исчерченность которой также обусловлена наличием разных типов миофиламентов. Миокард состоит из волокон, которые связаны между собой и формируют сетку. Эти волокна включают одно или двухъядерные клетки, что расположены в виде цепочки. Они получили название сократительных кардиомиоцитов.
Сократительные кардиомиоциты длиной от 50 до 120 микрометров, шириной - до 20 мкм. Ядро здесь располагается в центре цитоплазмы, в отличие от ядер поперечно полосатых волокон. Кардиомиоциты имеют больше саркоплазма и меньше миофибрилл, в сравнении со скелетными мышцами. В клетках сердечной мышцы находится много митохондрий, так как непрерывные сердечные сокращения требуют много энергии.
Вторая разновидность клеток миокарда - это проводящие кардиомиоциты, которые формируют проводящую систему сердца. Проводящие миоциты обеспечивают передачу импульса к сократительным мышечным клеткам.
Функции сердечной мышечной ткани:
- Насосная;
- обеспечивает ток крови в кровеносном русле.
Компоненты сократительной системы
Особенности строения мышечной ткани обусловлены выполняемыми функциями, возможностью принимать и проводить импульсы, способностью к сокращению. Механизм сокращения заключается в согласованной работе ряда элементов: миофибрилл, сократительных белков, митохондрий, миоглобина.
В цитоплазме мышечных клеток имеются особые сократительные нити - миофибриллы, сокращение которых возможно при содружественной работе белков - актина и миозина, а также при участии ионов Са. Митохондрии снабжают все процессы энергией. Также энергетические запасы образуют гликоген и липиды. Миоглобин необходим для связывания O 2 и формирование его запаса на период сокращения мышцы, так как во время сокращения идет сдавление кровеносных сосудов и снабжение мышц O 2 резко снижается.
Таблица. Соответствие между характеристикой мышечной ткани и ее видом
| Вид ткани | Характеристика |
|---|---|
| Гладкомышечная | Входит в состав стенок кровеносных сосудов |
| Структурная единица – гладкий миоцит | |
| Сокращается медленно, неосознанно | |
| Поперечная исчерченность отсутствует | |
| Скелетная | Структурная единица – многоядерное мышечное волокно |
| Свойственна поперечная исчерченность | |
| Сокращается быстро, осознанно |
Где находится мышечная ткань?
Гладкие мышцы являются составной частью стенок внутренних органов: желудочно-кишечного тракта, мочеполовой системы, сосудов. Входят в состав капсулы селезенки, кожных покровов, сфинктера зрачка.
Скелетная мускулатуразанимают около 40% от массы тела человека, с помощью сухожилий крепятся к костям. Из этой ткани состоят скелетные мышцы, мышцы рта, языка, глотки, гортани, верхнего участка пищевода, диафрагмы, мимическая мускулатура. Также поперечно полосатые мышцы находится в миокарде.
Чем мышечное волокно скелетной мышцы отличается от гладкой мышечной ткани?
Волокна поперечнополосатых мышц намного длиннее (до 12см), чем клеточные элементы гладкомышечной ткани (0,05-0,4мм). Также скелетные волокна имеют поперечную исчерченность благодаря особому расположению нитей актина и миозина. Для гладких мышц это не характерно.
В мышечных волокнах находится много ядер, а сокращение волокон сильное, быстрое и осознанное. В отличие от гладких мышц, клетки гладкомышечной ткани одноядерные, способны сокращаться в медленном темпе и неосознанно.
Выполняют очень важную функцию в организмах живых существ - формируют и выстилают все органы и их системы. Особое значение среди них имеет именно мышечная, так как ее значение в формировании наружной и внутренней полости всех структурных частей тела приоритетная. В данной статье рассмотрим, что собой представляет гладкая мышечная ткань, особенности строения ее, свойства.
Разновидности данных тканей
В составе животного организма имеется немного типов мышц:
- поперечно полосатая;
- гладкая мышечная ткань.
Обе они имеют свои характеристические черты строения, выполняемые функции и проявляемые свойства. Кроме того, их легко различить между собой. Ведь и та и другая имеют свой неповторимый рисунок, формирующийся благодаря входящим в состав клеток белковым компонентам.
Поперечнополосатая также подразделяется на два основных вида:
- скелетная;
- сердечная.
Само название отражает основные области расположения в организме. Ее функции чрезвычайно важны, ведь именно эта мускулатура обеспечивает сокращение сердца, движение конечностей и всех остальных подвижных частей тела. Однако, и гладкая мускулатура не менее значима. В чем заключаются ее особенности, рассмотрим дальше.
В целом можно заметить, что только слаженная работа, которую выполняет гладкая и поперечнополосатая мышечные ткани, позволяет всему организму успешно функционировать. Поэтому определить более или менее значимую из них невозможно.
Гладкая особенности строения
Основные необычные черты рассматриваемой структуры заключаются в строении и составе ее клеток - миоцитов. Как и любая другая, эта ткань образована группой клеток, схожих по строению, свойствам, составу и выполняемым функциям. Общие особенности строения можно обозначить в нескольких пунктах.
- Каждая клетка окружена плотным сплетением соединительнотканных волокон, что выглядит, словно капсула.
- Каждая структурная единица плотно прилегает к другой, межклетники практически отсутствуют. Это позволяет всей ткани быть плотноупакованной, структурированной и прочной.
- В отличие от поперечнополосатой коллеги, данная структура может включать в свой состав неодинаковые по форме клетки.
Это, конечно, не вся характеристика, которую имеет Особенности строения, как уже оговаривалось, заключаются именно в самих миоцитах, их функционировании и составе. Поэтому ниже этот вопрос будет рассмотрен подробнее.

Миоциты гладкой мускулатуры
Миоциты имеют разную форму. В зависимости от локализации в том или ином органе, они могут быть:
- овальными;
- веретеновидными удлиненными;
- округлыми;
- отростчатыми.
Однако в любом случае общий состав их сходен. Они содержат такие органоиды, как:
- хорошо выраженные и функционирующие митохондрии;
- комплекс Гольджи;
- ядро, чаще вытянутое по форме;
- эндоплазматический ретикулум;
- лизосомы.
Естественно, и цитоплазма с обычными включениями также присутствует. Интересен факт, что миоциты гладкой мускулатуры снаружи покрыты не только плазмолеммой, но и мембраной (базальной). Это обеспечивает им дополнительную возможность для контакта друг с другом.
Эти места соприкосновения составляют особенности гладкой мышечной ткани. Места контактов именуются нексусами. Именно через них, а также через поры, которые в этих местах имеются в мембране, происходит передача импульсов между клетками, обмен информацией, молекулами воды и другими соединениями.
Есть еще одна необычная черта, которую имеет гладкая мышечная ткань. Особенности строения ее миоцитов в том, что не все из них имеют нервные окончания. Поэтому настолько важны нексусы. Чтобы ни одна клетка не осталась без иннервации, и импульс мог передаться через соседнюю структуру по ткани.
Существует два основных типа миоцитов.
- Секреторные. Их основная функция заключается в выработке и накоплении гранул гликогена, сохранении множества митохондрий, полисом и рибосомальных единиц. Свое название эти структуры получили из-за белков, содержащиеся в них. Это актиновые филаменты и сократительные фибриновые нити. Данные клетки чаще всего локализуются по периферии ткани.
- Гладкие Имеют вид веретеновидных удлиненных структур, содержащих овальное ядро, смещенное к середине клетки. Другое название лейомиоциты. Отличаются тем, что имеют более крупные размеры. Некоторые частицы маточного органа достигают 500 мкм! Это достаточно значительная цифра на фоне всех остальных клеток в организме, больше разве что яйцеклетка.
Функция гладких миоцитов состоит также в том, что они синтезируют следующие соединения:
- гликопротеиды;
- проколлаген;
- эластаны;
- межклеточное вещество;
- протеогликаны.
Совместное взаимодействие и слаженная работа обозначенных типов миоцитов, а также их организация обеспечивают строение гладкой мышечной ткани.

Происхождение данной мускулатуры
Источник образования данного типа мускулатуры в организме не один. выделяют три основных варианта происхождения. Именно этим и объясняется различия, которые имеет строение гладкой мышечной ткани.
- Мезенхимное происхождение. такое имеет большая часть гладких волокон. Именно из мезенхими образуются практически все ткани, выстилающие внутреннюю часть полых органов.
- Эпидермальное происхождение. Само название говорит о местах локализации - это все кожные железы и их протоки. Именно они образованы гладкими волокнами, имеющими такой вариант появления. Потовые, слюнные, молочные, слезные - все эти железы выделяют свой секрет, благодаря раздражению клеток миоэпителиоцитов - структурных частичек рассматриваемого органа.
- Нейральное происхождение. Такие волокна локализуются в одном определенном месте - это радужка, одна из оболочек глаза. Сокращение или расширение зрачка иннервируется и управляется именно этими клетками гладкой мускулатуры.
Несмотря на разное происхождение, внутренний состав и выполняемые свойства всех в рассматриваемой ткани остаются примерно одинаковыми.
Основные свойства данной ткани
Свойства гладкой мышечной ткани соответствуют таковым и для поперечнополосатой. В этом они едины. Это:
- проводимость;
- возбудимость;
- лабильность;
- сократимость.
При этом существует и одна достаточно специфичная особенность. Если поперечнополосатая скелетная мускулатура способна быстро сокращаться (это хорошо иллюстрирует дрожь в теле человека), то гладкая может долго удерживаться в сжатом состоянии. Кроме того, ее деятельность не подчиняется воле и разуму человека. Так как иннервирует ее
Очень важным свойством является способность к длительному медленному растяжению (сокращению) и такому же расслаблению. Так, на этом основана работа мочевого пузыря. Под действием биологической жидкости (ее наполнением) он способен растягиваться, а затем сокращаться. Стенки его выстланы именно гладкой мускулатурой.

Белки клеток
Миоциты рассматриваемой ткани содержат много разных соединений. Однако наиболее важными из них, обеспечивающими выполнение функций сокращения и расслабления, являются именно белковые молекулы. Из них здесь содержатся:
- миозиновые нити;
- актин;
- небулин;
- коннектин;
- тропомиозин.
Эти компоненты обычно располагаются в цитоплазме клеток изолированно друг от друга, не образуя скоплений. Однако в некоторых органах у животных формируются пучки или тяжи, именуемые миофибриллами.
Расположение в ткани этих пучков в основном продольное. Причем как миозиновых волокон, так и актиновых. В результате образуется целая сеть, в которой концы одних сплетаются с краями других белковых молекул. Это важно для быстрого и правильного сокращения всей ткани.
Само сокращение происходит так: в составе внутренней среды клетки есть пиноцитозные пузырьки, в которых обязательно содержатся ионы кальция. Когда поступает нервный импульс, говорящий о необходимости сокращения, этот пузырек подходит к фибрилле. В результате ион кальция раздражает актин и он продвигается глубже между нитями миозина. Это приводит к затрагиванию плазмалеммы и в результате миоцит сокращается.

Гладкая мышечная ткань: рисунок
Если говорить о поперечнополосатой ткани, то ее легко узнать по исчерченности. Но вот что касается рассматриваемой нами структуры, то такого не происходит. Почему гладкая мышечная ткань рисунок имеет совсем иной, нежели близкая ей соседка? Это объясняется наличием и расположением белковых компонентов в миоцитах. В составе гладкой мускулатуры нити миофибрилл разной природы локализуются хаотично, без определенного упорядоченного состояния.
Именно поэтому рисунок ткани просто отсутствует. В поперечнополосатой нити актина последовательно сменяются поперечным миозином. В результате возникает рисунок - исчерченность, благодаря которой ткань и получила свое название.
Под микроскопом гладкая ткань выглядит очень ровной и упорядоченной, благодаря плотно прилегающим друг к другу продольно расположенным вытянутым миоцитам.
Области пространственного расположения в организме
Гладкая мышечная ткань образует достаточно большое количество важных внутренних органов в животном теле. Так, ей образованы:
- кишечник;
- половые органы;
- кровеносные сосуды всех типов;
- железы;
- органы выделительной системы;
- дыхательные пути;
- части зрительного анализатора;
- органы пищеварительной системы.
Очевидно, что места локализации рассматриваемой ткани крайне разнообразны и важны. Кроме того, следует заметить, что такая мускулатура формирует в основном те органы, которые подвержены автоматии в управлении.

Способы восстановления
Гладкая мышечная ткань образует достаточно важные структуры, что иметь способность к регенерации. Поэтому для нее характерны два основных пути восстановления при повреждениях различного рода.
- Митотическое деление миоцитов до образования нужного количества ткани. Самый распространенный простой и быстрый способ регенерации. Так происходит восстановление внутренней части любого органа, образованного гладкой мускулатурой.
- Миофибробласты способны трансформироваться в миоциты гладкой ткани при необходимости. Это более сложный и редко встречаемый путь регенерации данной ткани.
Иннервация гладкой мускулатуры
Гладкая свои выполняет независимо от желания или нежелания живого существа. Это происходит оттого, что ее иннервацию осуществляет вегетативная нервная система, а также отростки нервов ганглиев (спинальных).
Примером этому и доказательством может служить сокращение или увеличение размеров желудка, печени, селезенки, растяжение и сокращение мочевого пузыря.

Функции гладкой мышечной ткани
Каково же значение этой структуры? Зачем нужна ее следующие:
- длительное сокращение стенок органов;
- выработка секретов;
- способность отвечать на раздражения и воздействия возбудимостью.
Мышечные ткани объединяет способность к сокращению.
Особенности строения: сократительный аппарат, занимающий значительную часть в цитоплазме структурных элементов мышечной ткани и состоящий из актиновых и миозиновых филаментов, которые формируют органеллы специального назначения –миофибриллы .
Классификация мышечных тканей
1. Морфофункциональная классификация:
1) Поперечнополосатая, или исчерченная мышечная ткань: скелетная и сердечная;
2) Неисчерченная мышечная ткань: гладкая.
2. Гистогенетическая классификация (в зависимости от источников развития):
1) Соматического типа (из миотомов сомитов) – скелетная мышечная ткань (поперечнополосатая);
2) Целомического типа (из миоэпикардиальной пластинки висцерального листка спланхнотома) – сердечная мышечная ткань (поперечнополосатая);
3) Мезенхимного типа (развивается из мезенхимы) – гладкая мышечная ткань;
4) Из кожной эктодермы и прехордальной пластинки – миоэпителиальные клетки желёз (гладкие миоциты);
5) Нейрального происхождения (из нервной трубки) – мионейральные клетки (гладкие мышцы, суживающие и расширяющие зрачок).
Функции мышечной ткани : перемещение тела или его частей в пространстве.
СКЕЛЕТНАЯ МЫШЕЧНАЯ ТКАНЬ
Исчерченная (поперечно-полосатая) мышечная ткань составляет до 40% массы взрослого человека, входит в состав скелетных мышц, мышц языка, гортани и др. Относятся к произвольным мышцам, поскольку их сокращения подчиняются воле человека. Именно эти мышцы задействованы при занятии спортом.
Гистогенез. Скелетная мышечная ткань развивается из клеток миотомов миобластов. Различают головные, шейные, грудные, поясничные, крестцовые миотомы. Они разрастаются в дорзальном и вентральном направлениях. В них рано врастают ветви спинномозговых нервов. Часть миобластов дифференцируется на месте (образуют аутохтонную мускулатуру), а другие с 3 недели внутриутробного развития мигрируют в мезенхиму и, сливаясь друг с другом, образуют мышечные трубки (миотубы ) с крупными центрально ориентированными ядрами. В миотубах происходит дифференцировка специальных органелл миофибрилл. Первоначально они располагаются под плазмолеммой, а затем заполняют большую часть миотубы. Ядра смещаются к периферии. Клеточные центры и микротрубочки исчезают, грЭПС значительно редуцируется. Такая многоядерная структура называется симпласт , а для мышечной ткани – миосимпласт . Часть миобластов дифференцируется в миосателлитоциты, которые располагаются на поверхности миосимпластов и впоследствии принимают участие в регенерации мышечной ткани.
Строение скелетной мышечной ткани
Рассмотрим строение мышечной ткани на нескольких уровнях организации живого: на органном уровне (мышца как орган), на тканевом (непосредственно мышечная ткань), на клеточном (строение мышечного волокна), на субклеточном (строение миофибриллы) и на молекулярном уровне (строение актиновых и миозиновых нитей).
 На каритнке:
На каритнке:
1 — мышца икроножная (органный уровень), 2 — поперечный срез мышцы (тканевой уровень) — мышечные волокна, между которыми РВСТ: 3 — эндомизий, 4 — нервное волокно, 5 — кровеносный сосуд; 6 — поперечный срез мышечного волокна (клеточный уровень): 7 — ядра мышечного волокна — симпласта, 8 — митохондрия между миофибриллами, синим цветом — саркоплазматический ретикулум; 9 — поперечный срез миофибриллы (субклеточный уровень): 10 — тонкие актиновые нити, 11 — толстые миозиновые нити, 12 — головки толстых миозиновых нитей.
1) Органный уровень: строение мышцы как органа.
Скелетная мышца состоит из пучков мышечных волокон, связанных воедино системой соединительнотканных компонентов. Эндомизий – прослойки РВСТ между мышечными волокнами, где проходят кровеносные сосуды, нервные окончания. Перимизий – окружает 10-100 пучков мышечных волокон. Эпимизий – наружная оболочка мышцы, представлена плотной волокнистой тканью.
2) Тканевой уровень: строение мышечной ткани.
 Структурно-функциональной единицей скелетной поперечнополосатой (исчерченной) мышечной ткани является мышечное волокно
– цилиндрической формы образование диаметром 50 мкм и длиной от 1 до 10-20 см. Мышечное волокно состоит из 1) миосимпласта
(образование его смотри выше, строение – ниже), 2) мелких камбиальных клеток – миосателлитоцитов
, прилежащих к поверхности миосимпласта и располагающиеся в углублениях его плазмолеммы, 3) базальной мембраны, которой покрыта плазмолемма. Комплекс плазмолеммы и базальной мембраны называется сарколемма
. Для мышечного волокна характерна поперечная исчерченность, ядра смещены на периферию. Между мышечными волокнами – прослойки РВСТ (эндомизий).
Структурно-функциональной единицей скелетной поперечнополосатой (исчерченной) мышечной ткани является мышечное волокно
– цилиндрической формы образование диаметром 50 мкм и длиной от 1 до 10-20 см. Мышечное волокно состоит из 1) миосимпласта
(образование его смотри выше, строение – ниже), 2) мелких камбиальных клеток – миосателлитоцитов
, прилежащих к поверхности миосимпласта и располагающиеся в углублениях его плазмолеммы, 3) базальной мембраны, которой покрыта плазмолемма. Комплекс плазмолеммы и базальной мембраны называется сарколемма
. Для мышечного волокна характерна поперечная исчерченность, ядра смещены на периферию. Между мышечными волокнами – прослойки РВСТ (эндомизий).
3) Клеточный уровень: строение мышечного волокна (миосимпласта).
Термин «мышечное волокно» подразумевает «миосимпласт», поскольку миосимпласт обеспечивает функцию сокращения, миосателлитоциты участвуют только в регенерации.
Миосимпласт , как и клетка, состоит из 3-х компонентов: ядра (точнее множества ядер), цитоплазмы (саркоплазма) и плазмолеммы (которая покрыта базальной мембраной и называется сарколемма). Почти весь объём цитоплазмы заполнен миофибриллами – органеллами специального назначения, органеллы общего назначения: грЭПС, аЭПС, митохондрии, комплекс Гольджи, лизосомы, а также ядра смещены на периферию волокна.
В мышечном волокне (миосимпласте) различают функциональные аппараты: мембранный , фибриллярный (сократительный) и трофический .
Трофический аппарат включает ядра, саркоплазму и цитоплазматические органеллы: митохондрии (синтез энергии), грЭПС и комплекс Гольджи (синтез белков – структурных компонентов миофибрилл), лизосомы (фагоцитоз изношенных структурных компонентов волокна).
Мембранный аппарат : каждое мышечное волокно покрыто сарколеммой, где различают наружную базальную мембрану и плазмолемму (под базальной мембраной), которая образует впячивания (Т -трубочки). К каждой Т -трубочке примыкают по две цистерны триаду : две L -трубочки (цистерны аЭПС) и одна Т -трубочка (впячивание плазмолеммы). В цистернах аЭПС концентрируются Са 2+ , необходимый при сокращении. К плазмолемме снаружи прилежат миосателлитоциты. При повреждении базальной мембраны запускается митотический цикл миосателлитоцитов.
Фибриллярный аппарат .Большую часть цитоплазмы исчерченных волокон занимают органеллы специального назначения – миофибриллы, ориентированы продольно, обеспечивающие сократительную функцию ткани.
4) Субклеточный уровень: строение миофибриллы.
При исследовании мышечных волокон и миофибрилл под световым микроскопом, отмечается чередование в них темных и светлых участков – дисков. Темные диски отличаются двойным лучепреломлением и называются анизотропными дисками, или А - дисками. Светлые диски не обладают двойным лучепреломлением и называются изотропными, или I -дисками.
В середине диска А имеется более светлый участок – Н -зона, где содержатся только толстые нити белка миозина. В середине Н -зоны (значит и А -диска) выделяется более темная М -линия, состоящая из миомезина (необходим для сборки толстых нитей и их фиксации при сокращении). В середине диска I расположена плотная линия Z , которая построена из белковых фибриллярных молекул. Z -линия соединена с соседними миофибриллами с помощью белка десмина, и поэтому все названные линии и диски соседних миофибрилл совпадают и создается картина поперечнополосатой исчерченности мышечного волокна.
Структурной единицей миофибриллы является саркомер (S ) — это пучок миофиламентов заключенный между двумя Z -линиями. Миофибрилла состоит из множества саркомеров. Формула, описывающая структуру саркомера:
S = Z 1 + 1/2 I 1 + А + 1/2 I 2 + Z 2

5) Молекулярный уровень: строение актиновых и миозиновых филаментов .
Под электронным микроскопом миофибриллы представляют агрегаты из толстых, или миозиновых , и тонких, или актиновых , филаментов. Между толстыми филаментами располагаются тонкие филаменты (диаметр 7-8 нм).
Толстые филаменты, или миозиновые нити, (диаметр 14 нм, длина 1500 нм, расстояние между ними 20-30 нм) состоят из молекул белка миозина, являющимся важнейшим сократительным белком мышцы, по 300-400 молекул миозина в каждой нити. Молекула миозина – это гексамер, состоящий из двух тяжелых и четырех легких цепей. Тяжелые цепи представляют собой две спирально закрученные полипептидные нити. Они несут на своих концах шаровидные головки. Между головкой и тяжелой цепью находится шарнирный участок, с помощью которого головка может изменять свою конфигурацию. В области головок – легкие цепи (по две на каждой). Молекулы миозина уложены в толстой нити таким образом, что их головки обращены наружу, выступая над поверхностью толстой нити, а тяжелые цепи образуют стержень толстой нити.
Миозин обладает АТФ-азной активностью: высвобождающаяся энергия используется для мышечного сокращения.
Тонкие филаменты, или актиновые нити, (диаметр 7-8 нм), образованы тремя белками: актином, тропонином и тропомиозином. Основным по массе белком является актин, который образует спираль. Молекулы тропомиозина располагаются в желобке этой спирали, молекулы тропонина располагаются вдоль спирали.
Толстые нити занимают центральную часть саркомера – А -диск, тонкие занимают I - диски и частично входят между толстыми миофиламентами. Н -зона состоит только из толстых нитей.
В покое взаимодействие тонких и толстых нитей (миофиламентов) невозможно, т.к. миозин-связывающие участки актина заблокированы тропонином и тропомиозином. При высокой концентрации ионов кальция конформационные изменения тропомиозина приводят к разблокированию миозин-связывающих участков молекул актина.
Двигательная иннервация мышечного волокна . Каждое мышечное волокно имеет собственный аппарат иннервации (моторная бляшка) и окружено сетью гемокапилляров, располагающихся в прилежащей РВСТ. Этот комплекс называется мион. Группа мышечных волокон, которые иннервируются одним мотонейроном, называется нервно-мышечной единицей. Мышечные волокна в этом случае могут располагаться не рядом (одно нервное окончание может контролировать от одного до десятков мышечных волокон).
При поступлении нервных импульсов по аксонам двигательных нейронов происходит сокращение мышечного волокна .
Сокращение мышцы
При сокращении мышечные волокна укорачиваются, но длина актиновых и миозиновых филаментов в миофибриллах не изменяется, а происходит их движение друг относительно друга: миозиновые нити вдвигаются в пространства между актиновыми а, актиновые – между миозиновыми. В результате этого уменьшается ширина I -диска, H -полоски и уменьшается длина саркомера; ширина А -диска не изменяется.
Формула саркомера при полном сокращении:S = Z 1 + А + Z 2
Молекулярный механизм мышечного сокращения
1. Прохождение нервного импульса через нервно-мышечный синапс и деполяризация плазмолеммы мышечного волокна;
2. Волна деполяризации проходит по Т -трубочкам (впячивания плазмолеммы) до L -трубочек (цистерны саркоплазматического ретикулума);
3. Открытие кальциевых каналов в саркоплазматическом ретикулуме и выход ионов Са 2+ в саркоплазму;
4. Кальций диффундирует к тонким нитям саркомера, связывается с тропонином С, приводя к конформационным изменениям тропомиозина и освобождая активные центры для связывания миозина и актина;
5. Взаимодействие миозиновых головок с активными центрами на молекуле актина с образованием актино-миозиновых «мостиков»;
6. Миозиновые головки «шагают» по актину, образуя в ходе перемещения новые связи актина и миозина, при этом актиновые нити подтягиваются в пространство между миозиновыми нитями к M -линии, сближая две Z -линии;
7. Расслабление: Са 2+ -АТФ-аза саркоплазматического ретикулума закачивает Са 2+ из саркоплазмы в цистерны. В саркоплазме концентрация Са 2+ становится низкой. Разрываются связи тропонина С с кальцием, тропомиозин закрывает миозин-связывающие участки тонких нитей и препятствует их взаимодействию с миозином.
Каждое движение головки миозина (присоединение к актину и отсоединение) сопровождается затратой энергии АТФ.
Чувствительная иннервация (нервно-мышечные веретена). Интрафузальные мышечные волокна вместе с чувствительными нервными окончаниями формируют нервно-мышечные веретена, являющиеся рецепторами скелетной мышцы. Снаружи сформирована капсула веретена. При сокращении поперечно-полосатых (исчерченных) мышечных волокон изменяется натяжение соединительно-тканной капсулы веретена и соответственно изменяется тонус интрафузальных (расположенных под капсулой) мышечных волокон. Формируется нервный импульс. При избыточном растяжении мышцы возникает чувство боли.
Классификация и типы мышечных волокон
1. По характеру сокращения: фазные и тонические мышечные волокна. Фазные способны осуществлять быстрые сокращения, но не могут длительно удерживать достигнутый уровень укорочения. Тонические мышечные волокна (медленные) обеспечивают поддержание статического напряжения или тонуса, что играет роль в сохранения определённого положения тела в пространстве.
2. По биохимическим особенностям и цвету выделяют красные и белые мышечные волокна . Цвет мышцы обусловлен степенью васкуляризации и содержанием миоглобина. Характерной особенностью красных мышечных волокон является наличие многочисленных митохондрий, цепи которых располагаются между миофибриллами. В белых мышечных волокнах митохондрий меньше и они располагаются равномерно в саркоплазме мышечного волокна.
3. По типу окислительного обмена : оксидативные, гликолитические и промежуточные . Идентификация мышечных волокон основана на выявлении активности фермента сукцинатдегидрогеназы (СДГ), которая является маркером для митохондрий и цикла Кребса. Активность этого фермента свидетельствует о напряженности энергетического метаболизма. Выделяют мышечные волокна А -типа (гликолитические) с низкой активностью СДГ, С -тип (оксидативные) с высокой активностью СДГ. Мышечные волокна В -типа занимают промежуточное положение. Переход мышечных волокон от А -типа в С -тип маркирует изменения от анаэробного гликолиза к метаболизму, зависящему от кислорода.
У спринтеров (спортсменов, когда нужен быстрое недолгое сокращение, культуристов) тренировки и питание направлено на развитие гликолитических, быстрых, белых мышечных волкон : в них много запасов гликогена и энергия добывается преимущественно анаэолбным путём (белое мясо у курицы). У стайеров (спортсменов — марафонцев, в тех видах спорта, где необходима выносливость) преобладают оксидативные, медленные, красные волокна в мышцах — в них много митохондрий для аэробного гликолиза, кровеносных сосудов (нужен кислород).
4. В исчерченных мышцах различают два вида мышечных волокон: экстрафузальные , которые преобладают и обуславливают собственно сократительную функцию мышцы и интрафузальные , входящие в состав проприоцепторов – нервно-мышечных веретен.
Факторами, определяющими структуру и функцию скелетной мышцы являются влияние нервной ткани, гормональное влияние, местоположение мышцы, уровень васкуляризации и двигательной активности.
СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ
Сердечная мышечная тканьнаходится в мышечной оболочке сердца (миокард) и в устьях связанных с ним крупных сосудов. Имеет клеточный тип строения и основным функциональным свойством служит способность к спонтанным ритмическим сокращениям (непроизвольные сокращения).
Развивается из миоэпикардиальной пластинки (висцеральный листок спланхнотома мезодермы в шейном отделе), клетки которой размножаются митозом, а потом дифференцируются. В клетках появляются миофиламенты, которые далее формируют миофибриллы.
Строение . Структурная единица сердечной мышечной ткани – клетка кардиомиоцит. Между клетками находятся прослойки РВСТ с кровеносными сосудами и нервами.
Типы кардиомиоцитов : 1) типичные (рабочие, сократительные), 2) атипичные (проводящие), 3) секреторные .
Типичные кардиомиоциты
Типичные (рабочие, сократительные) кардиомиоциты – клетки цилиндрической формы, длиной до 100-150 мкм и диаметром 10-20 мкм. Кардиомиоциты образуют основную часть миокарда, соединены друг с другом в цепочки основаниями цилиндров. Эти зоны называют вставочными дисками , в которых выделяют десмосомальные контакты и нексусы (щелевидные контакты). Десмосомы обеспечивают механическое сцепление, которое препятствует расхождению кардиомиоцитов. Щелевидные контакты способствуют передаче сокращения от одного кардиомиоцита к другому.
Каждый кардиомиоцит содержат одно или два ядра, саркоплазму и плазмолемму, окружённую базальной мембраной. Различают функциональные аппараты, такие же, как в мышечном волокне: мембранный , фибриллярный (сократительный), трофический, а также энергетический .
Трофический аппарат включает ядро, саркоплазму и цитоплазматические органеллы: грЭПС и комплекс Гольджи (синтез белков – структурных компонентов миофибрилл), лизосомы (фагоцитоз структурных компонентов клетки). Кардиомиоциты, как и олокна скелетной мышечной ткани, характеризуются наличием в их саркоплазме железосодержащего кислород-связывающего пигмента миоглобина, придающего им красный цвет и сходного по строению и функции с гемоглобином эритроцитов.
Энергетический аппарат представлен митохондриями и включениями, расщепление которых обеспечивает получение энергии. Митохондрии многочисленны, лежат рядами между фибриллами, у полюсов ядра и под сарколеммой. Энергия, необходимая кардиомиоцитам, получается путём расщепления: 1) основного энергетического субстрата этих клеток – жирных кислот , которые депонируются в виде триглицеридов в липидных каплях; 2) гликогена, находящегося в гранулах, расположенных между фибриллами.
Мембранный аппарат : каждая клетка покрыта оболочкой, состоящей из комплекса плазмолеммы и базальной мембраны. Оболочка образует впячивания (Т -трубочки). К каждой Т -трубочке примыкает одна цистерна (в отличие от мышечного волокна – там 2 цистерны) саркоплазматического ретикулума (видоизменённая аЭПС), образуя диаду : одна L -трубочка (цистерна аЭПС) и одна Т -трубочка (впячивание плазмолеммы). В цистернах аЭПС ионы Са 2+ накапливаются не так активно, как в мышечных волокнах.
Фибриллярный (сократительный) аппарат .Большую часть цитоплазмы кардиомиоцита занимают органеллы специального назначения – миофибриллы, ориентированы продольно и расположенные по периферии клетки.Сократительный аппарат рабочих кардиомиоцитовсходен со скелетными мышечными волокнами. При расслаблении, ионы кальция выделяются в саркоплазму с низкой скоростью, что обеспечивает автоматизм и частые сокращения кардиомиоцитов. Т -трубочки широкие и образуют диады (одна Т -трубочка и одна цистерна сети), которые сходятся в области Z -линии.
Кардиомиоциты, связываясь с помощью вставочных дисков, образуют сократительные комплексы, которые способствуют синхронизации сокращения, между кардиомиоцитами соседних сократительных комплексов образуются боковые анастомозы.
Функция типичных кардиомиоцитов : обеспечение силы сокращения сердечной мышцы.
Проводящие (атипичные) кардиомиоциты обладают способностью к генерации и быстрому проведению электрических импульсов. Они образуют узлы и пучки проводящей системы сердца и разделяются на несколько подтипов: пейсмекеры (в синоатриальном узле), переходные (в атрио-вентрикулярном узле) и клетки пучка Гиса и волокон Пуркинье. Проводящие кардиомиоциты характеризуются слабым развитием сократительного аппарата, светлой цитоплазмой и крупными ядрами. В клетках нет Т-трубочек и поперечной исчерченности, поскольку миофибриллы расположены неупорядоченно.
Функция атипичных кардиомиоцитов – генерация импульсов и передача на рабочие кардиомиоциты, обеспечивая автоматизм сокращения миокарда.
Секреторные кардиомиоциты
Секреторные кардиомиоцитынаходятся в предсердиях, преимущественно в правом; характеризуются отростчатой формой и слабым развитием сократительного аппарата. В цитоплзме, вблизи полюсов ядра – секреторные гранулы, содержащие натриуретический фактор, или атриопептин (гормон, регулирующий артериальное давление). Гормон вызывает потерю натрия и воды с мочой, расширение сосудов, снижение давления, угнетение секреции альдостерона, кортизола, вазопрессина.
Функция секреторных кардиомиоцитов : эндокринная.
Регенерация кардиомиоцитов. Для кардиомиоцитов характерна только внутриклеточная регенерация. Кардиомиоциты не способны к делению, у них отсутствуют камбиальные клетки.
ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ
Гладкая мышечная ткань образует стенки внутренних полых органов, сосудов; характеризуется отсутствием исчерченности, непроизвольными сокращениями. Иннервация осуществляется вегетативной нервной системой.
Структурно-функциональная единица неисчерченной гладкой мышечной ткани – гладкая мышечная клетка (ГМК), или гладкий миоцит. Клетки имеют веретенообразную форму длиной 20-1000 мкм и толщиной от 2 до 20 мкм. В матке клетки имеют вытянутую отростчатую форму.
Гладкий миоцит
Гладкий миоцит состоит из расположенного в центре ядра палочковидной формы, цитоплазмы с органеллами и сарколеммы (комплекс плазмолеммы и базальной мембраны). В цитоплазме у полюсов находится комплекс Гольджи, много митохондрий, рибосом, развит саркоплазматический ретикулум. Миофиламенты расположены косо или вдоль продольной оси. В ГМК актиновые и миозиновые филаменты не формируют миофибрилл. Актиновых нитей больше и они прикрепляются к плотным тельцам, которые образованы специальными сшивающими белками. Рядом с актиновыми нитями располагаются мономеры миозина (микромиозин). Обладая разной длиной, они значительно короче тонких нитей.
Сокращение гладких мышечных клеток осуществляется при взаимодействии актиновых филаментов и миозина. Сигнал, идущий по нервным волокнам, обуславливает выделение медиатора, что изменяет состояние плазмолеммы. Она образует колбовидные впячивания (кавеолы), где концентрируются ионы кальция. Сокращение ГМК индуцируется притоком ионов кальция в цитооплазму: кавеолы отшнуровываются и вместе с ионами кальция попадают в клетку. Это приводит к полимеризации миозина и взаимодействию его с актином. Актиновые нити и плотные тельца сближаются, усилие передается на сарколемму и ГМК укорачивается. Миозин в гладких миоцитах способен взаимодействовать с актином только после фосфорилирования его легких цепей особым ферментом – киназой легких цепей. После прекращения сигнала ионы кальция покидают кавеолы; миозин деполяризуется, теряет сродство к актину. В результате комплексы миофиламентов распадаются; сокращение прекращается.
Особые типы мышечных клеток
Миоэпителиальные клетки являются производными эктодермы, не имеют исчерченности. Окружают секреторные отделы и выводные протоки желез (слюнных, молочных, слезных). С железистыми клетками они связаны десмосомами. Сокращаясь, способствуют выделению секрета. В концевых (секреторных) отделах форма клеток отросчатая, звездчатая. Ядро в центре, в цитоплазме, преимущественно в отростках локализованы миофиламенты, которые образуют сократительный аппарат. В этих клетках есть и цитокератиновые промежуточные филаменты, что подчеркивает их сходство с эпителиоцитами.
Мионейральные клетки развиваются из клеток наружного слоя глазного бокала и образуют мышцу, суживающую зрачок и мышцу, расширяющую зрачок. По строению первая мышца сходна с ГМК мезенхимного происхождения. Мышца, расширяющая зрачок образована отростками клеток, располагающимися радиально, а ядросодержащая часть клетки находится между пигментным эпителием и стромой радужки.
Миофибробласты относятся к рыхлой соединительной ткани и представляют собой видоизмененные фибробласты. Они проявляют свойства фибробластов (синтезируют межклеточное вещество) и гладких миоцитов (обладают выраженными сократительными свойствами). Как вариант этих клеток можно рассматривать миоидные клетки в составе стенки извитого семенного канальца яичка и наружного слоя теки фолликула яичника. При заживлении раны часть фибробластов синтезирует гладкомышечные актины и миозины. Миофибробласты обеспечивают стягивание краёв раны.
Эндокринные гладкие миоциты – это видоизмененные ГМК, представляющие основной компонент юкстагломерулярного аппарата почек. Они находятся в стенке артериол почечного тельца, имеют хорошо развитый синтетический аппарат и редуцированный сократительный. Продуцируют фермент ренин, находящийся в гранулах и попадающий в кровь механизмом экзоцитоза.
Регенерация гладкой мышечной ткани. Гладкие миоциты характеризуются внутриклеточной регенерацией. При повышении функциональной нагрузки происходит гипертрофия миоцитов и в некоторых органах гиперплазия (клеточная регенерация). Так, при беременности гладко-мышечные клетки матки могут увеличиваться в 300 раз.